


非周期性脑电图是指脑电图(EEG)中不具有明显周期性的特征。
在脑电图中,正常情况下会观察到各种频率的波形和节律性活动,如α波、β波、δ波等,它们通常具有一定的周期性和规律性。但是,有些情况下脑电图可能出现不规则、不稳定的波形,即非周期性脑电图。
非周期性脑电图可能表现为以下特征:
关于大脑如何产生脑电图频谱趋势,突触时间尺度假说(the synaptic timescale hypothesis)预测,脑电图频谱趋势是突触电流呈指数衰减的自然结果,因此异步网络活动将产生具有以下特征的频谱:
尽管有这些假设,非周期性脑电图的概念本身仍然存在争议。
一些人认为,脑电图频谱的明显趋势是由于大脑节律较慢而招募更多神经元引起的附带现象。根据这一观点,脑电图频谱不需要去除趋势,频谱指数是大脑节律各种变化的综合测量。
因此,三个问题仍然悬而未决:
(1) 脑电图信号能否反映心律失常的神经活动?
(2) 如果是这样,这些信号如何形成脑电图谱?
(3) 脑电图谱是否需要去趋势,如果需要,**生理意义的去趋势方法是什么?
该研究将头皮电位的数值正演模型与单神经元偶极子的生物物理计算结合起来。通过这种方法,实验模拟了由表现出一系列动态的网络生成的生物物理真实脑电图信号。这些模拟揭示了除了大脑节律之外的几种影响脑电图信号并共同塑造频谱趋势的机制。
为了测试模型预测,实验记录了接受药物异丙酚输注的人类脑电图,这注射药物是一种针对 GABA 受体并减缓抑制性突触电流衰减时间的全身麻醉剂。这些实验确定了丙泊酚给药期间的特定脑电图变化,这些变化可能会影响脑节律估计。利用实验的建模见解,该研究纠正了这些污染源,并重新评估了意识丧失的已知EEG信号。
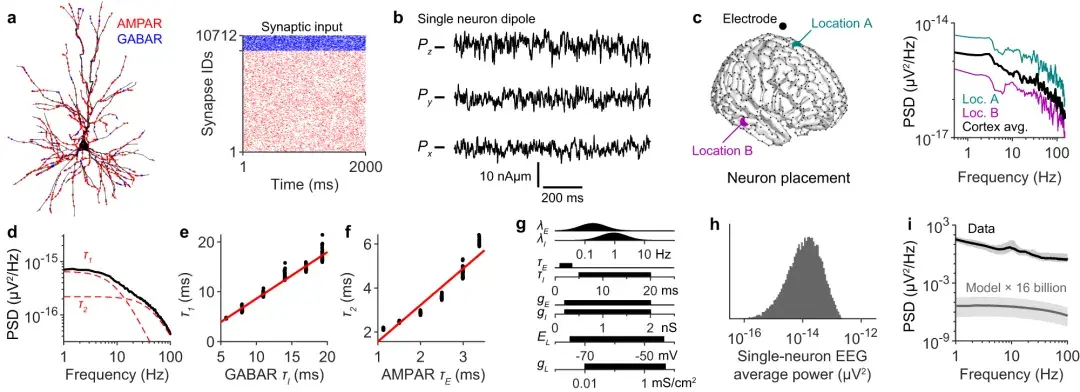
a :2/3 层锥体神经元的形态示例。 AMPAR(红色)和 GABAR(蓝色)突触的输入使用泊松尖峰序列进行模拟,如光栅图中所示。
b :单神经元偶极子向量的 x、y 和 z 分量,根据 ( a ) 计算得出。
c左 :在标记的电极位置模拟单神经元 EEG 信号,神经元位于不同的源位置,例如位置 A 和 B。
c右 :通过对源位置进行平均计算得到的位置平均 EEG 频谱(黑色)在大脑模板上以黑点显示。Loc.=位置;平均=平均。
d :由 11 个**性神经元形态的 1000 次模拟生成的位置平均频谱(表1)。频谱由方程拟合。
e :失活动力学的同时计算酉谱,并估计参数 。红线的斜率为 1。
f :与e相同,但显示作为 AMPAR 失活动力学的函数。
g :模型参数的采样分布。、和表示 AMPAR 突触的平均输入速率、失活时间常数和**电导。、和表示 GABAR 突触的相同参数。和表示无源膜漏电流的反转电位和电导。
h :单神经元 EEG 功率的分布(如c中的位置平均)基于 20,000 次模拟,参数从 ( g ) 中所示的分布中采样,采样的形态如表1中所述。
i :黑色:14 名受试者的中位脑电图频谱,误差带指示*小和**频谱密度。灰色:接收泊松突触输入(灰色)的 160 亿个不相关神经元的预测脑电图谱,参数值从 ( g )中的分布中采样。误差带反映 5–95% 的分位数范围。
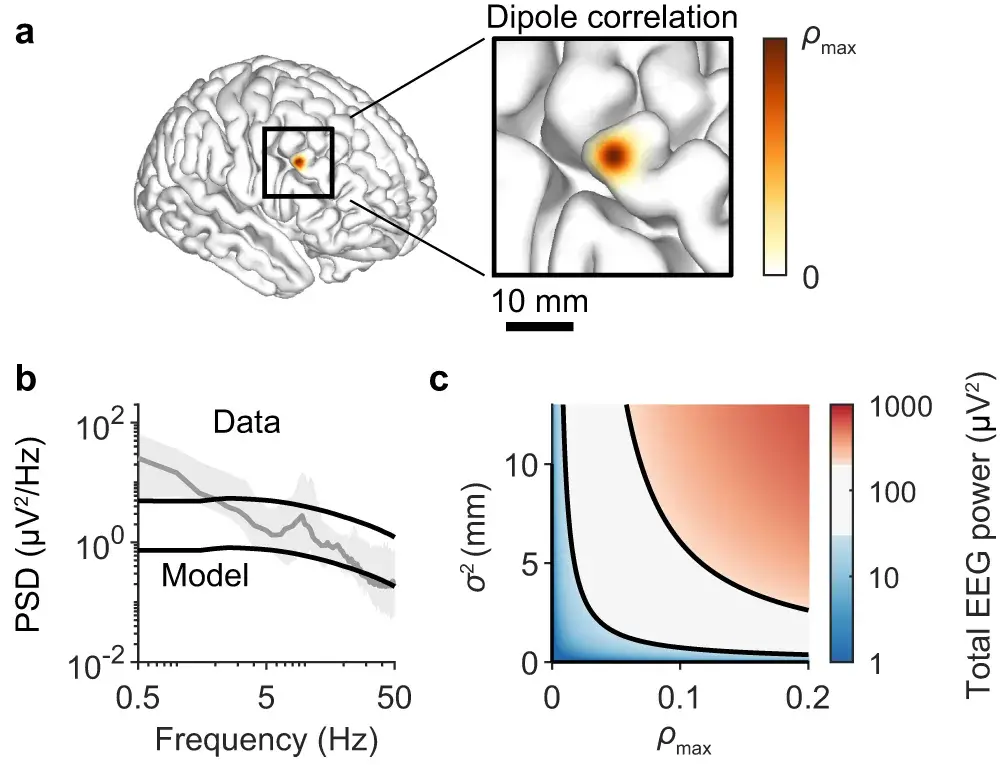
a :核是一个高斯函数,峰值为和方差。
b :14 名人类受试者的中位 EEG 频谱(灰色:与图 1i 相同的误差带)和按任意量缩放的模拟单一频谱(黑色:与图 1d相同)以确定频谱趋势幅度的下限和上限。
c :由 160 亿个具有偶极子的神经元与 (a)中的内核耦合产生的总模拟 EEG 功率热图和的各种值进行参数化。黑线是水平曲线,**(b)中获得的频谱趋势幅度的下限和上限。
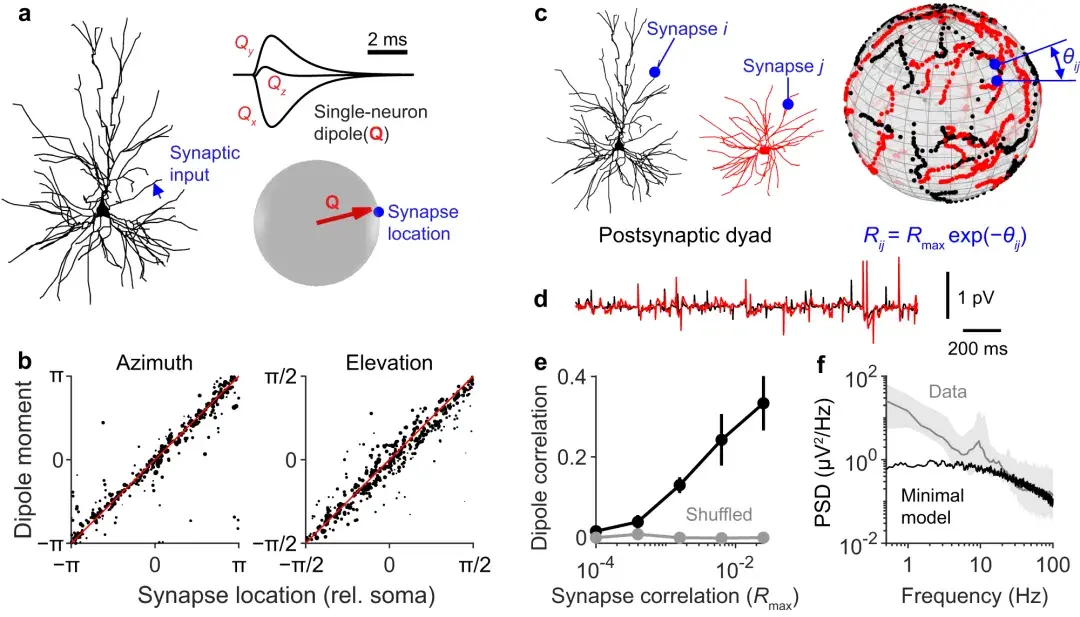
a :单个突触在蓝色箭头指定的位置被**,在单神经元偶极子Q中产生响应。响应峰值处的偶极矢量朝向突触位置,以体细胞为原点在球坐标中定义。
b :使用不同的神经元形态和突触位置重复a中的模拟600 次。绘制响应峰值处的偶极子方向与受刺激突触位置的关系,显示出很强的线性关系。
c :显示偶极相关性*小模型的示意图。二元体中每个突触后神经元的突触被投影到一个球体上。然后定义所有突触之间的相关矩阵,使得球体上以角度θij分隔的突触通过Rmaxexp(-θij)。
d :(c)中所示的两个神经元的单神经元 EEG 信号示例。e两个单神经元偶极子之间的相关性作为函数。垂直线**平均值的 95% 置信区间(n = 11 个不同的形态对)。当突触位置被打乱时,偶极子不再相关(灰色)。
f :使用*小模型(黑色)计算的接收输入的神经元的单一频谱,进行缩放以将频谱的形状与 14 名受试者的中位脑电图频谱进行比较(灰线;与图 1i 相同)。
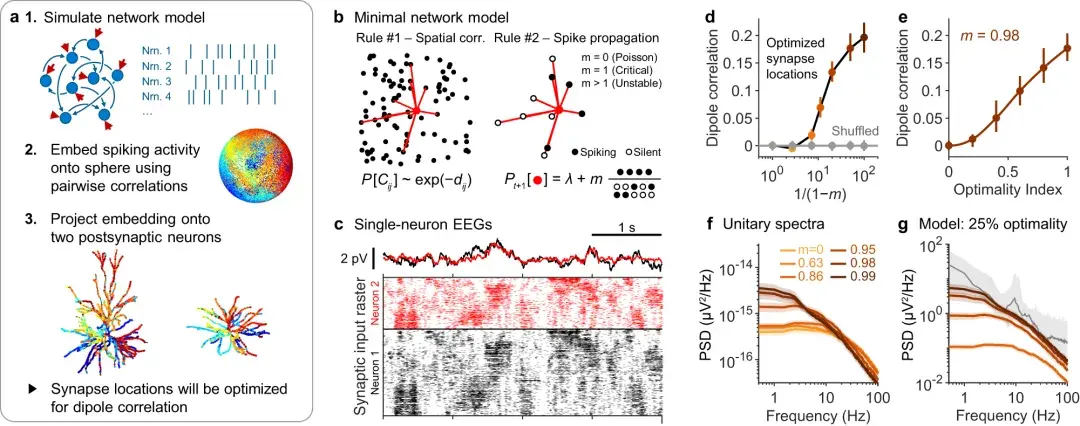
a :优化偶极子相干性算法的图示。
b :用于在突触前网络中产生时空同步的规则的图示。
规则#1:通过使成对连接的概率随距离呈指数下降来强制执行网络拓扑。
规则#2:神经元放电的概率取决于基线放电率加上其平均活动邻居按m缩放。m是网络的分支数。corr.=相关性。
c :当突触位置已按 ( a )中所述进行优化时,的网络模型的输入的单神经元 EEG 信号示例。底部:两个神经元突触输入的栅格图。
d :两个单神经元偶极子之间的相关性作为突触前网络分支数m的函数。
e :偶极子相关性可以通过将突触放置在次优位置来调整。模拟**性指数之间的三次插值用实线显示。n = 500 次模拟的中值(点)和 5–95% 分位数范围(垂直线)。
f :针对不同支化数计算的单一光谱。误差带反映平均值的 95% 置信区间。
g :来自 ( f ) 的单一光谱基于突触**性指数 0.25 进行缩放(颜色与f相同)的值由e 确定,*终的缩放因子如图2所示。灰色:14名受试者的中位脑电图谱(与图1i相同 )。
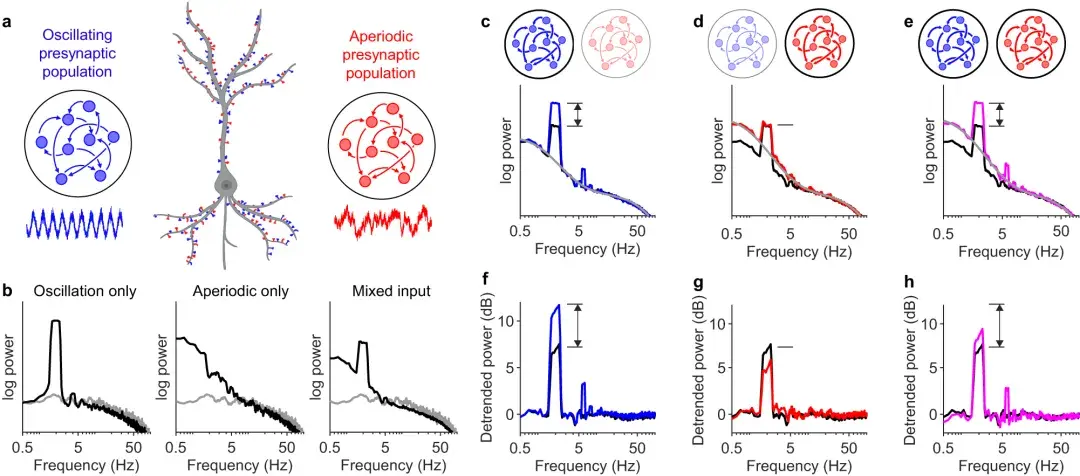
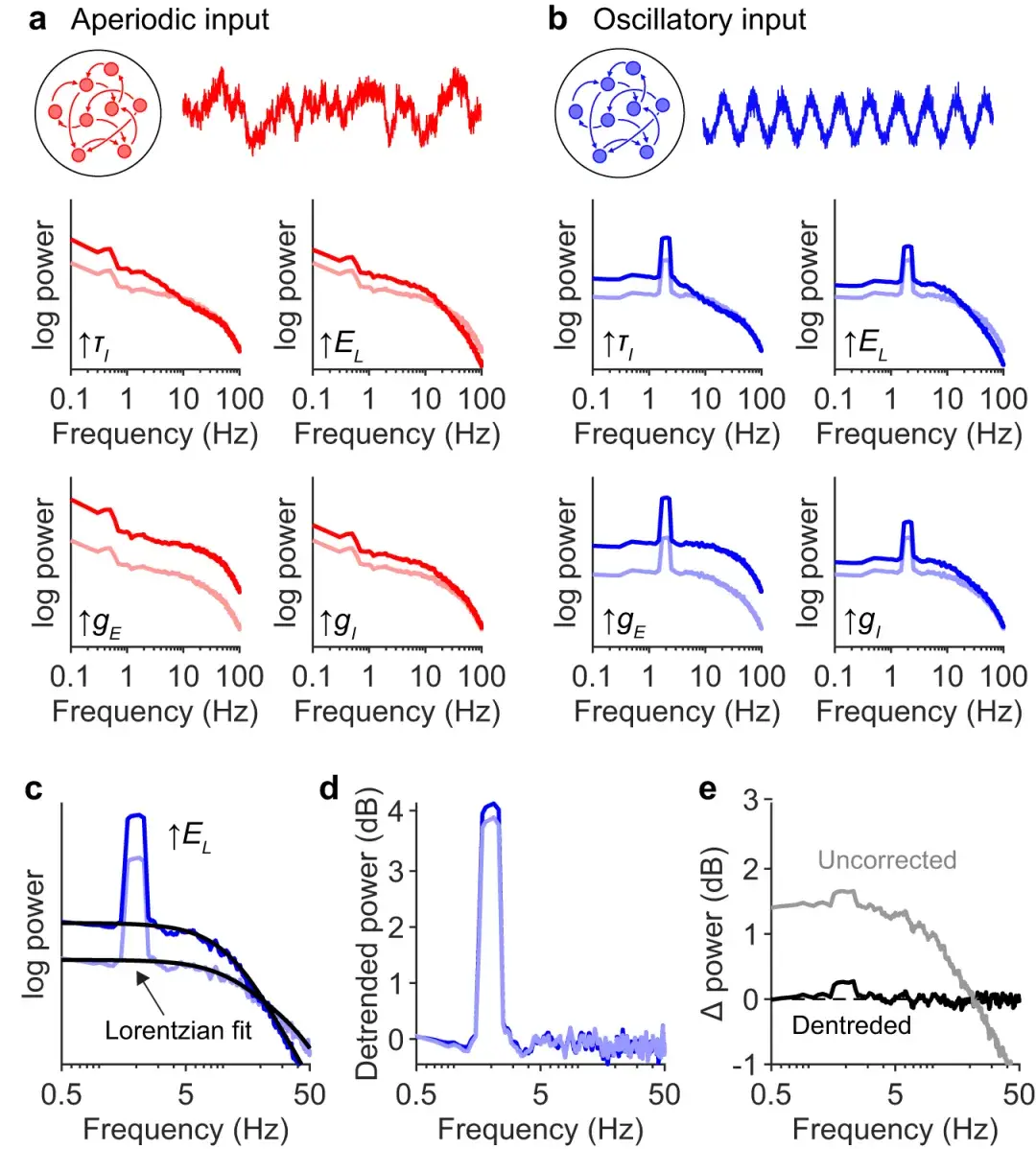
a :混合输入模型的图示:一半神经元突触接收振荡节律输入(蓝色),另一半接收来自亚临界网络(红色)的输入。
振荡和亚临界动力学的强度分别通过调节两个参数(αR和αA)来调节,这两个参数决定了突触输入与同质泊松过程的差异程度。
b :接收泊松输入的神经元的酉谱(灰色),与完全从振荡群体(左;黑色)、完全从亚临界群体(中;黑色)或混合输入接收输入的神经元的酉谱相比,如(a)(右;黑色)。
c :从 0.1(黑色)增加到 0.5(蓝色)来加强振荡输入固定为 0.1。使用类似 FOOOF 的算法拟合光谱,不同之处在于这里的非周期分量是用方程 1 建模的。
d :从 0.1(黑色)增加到 0.5(红色)来加强非周期输入固定为 0.1。
e :和从 0.1(黑色)增加到 0.5(洋红色),振荡输入和非周期输入都得到加强。
f :振荡强度增加之前(黑色)和之后(蓝色)接收混合输入的神经元的去趋势谱。( c )中的单一光谱除以灰色实线。
g :非周期性强度增加之前(黑色)和之后(红色)的去趋势功率。请注意,尽管振荡强度保持不变,但 2 Hz 处的去趋势功率却下降了。
h :振荡和非周期性强度增加之前(黑色)和之后(洋红色)的去趋势功率。请注意,尽管振荡强度增加了相同的量,但2 Hz 处的去趋势功率并未像 ( f)中那样增加那么多。
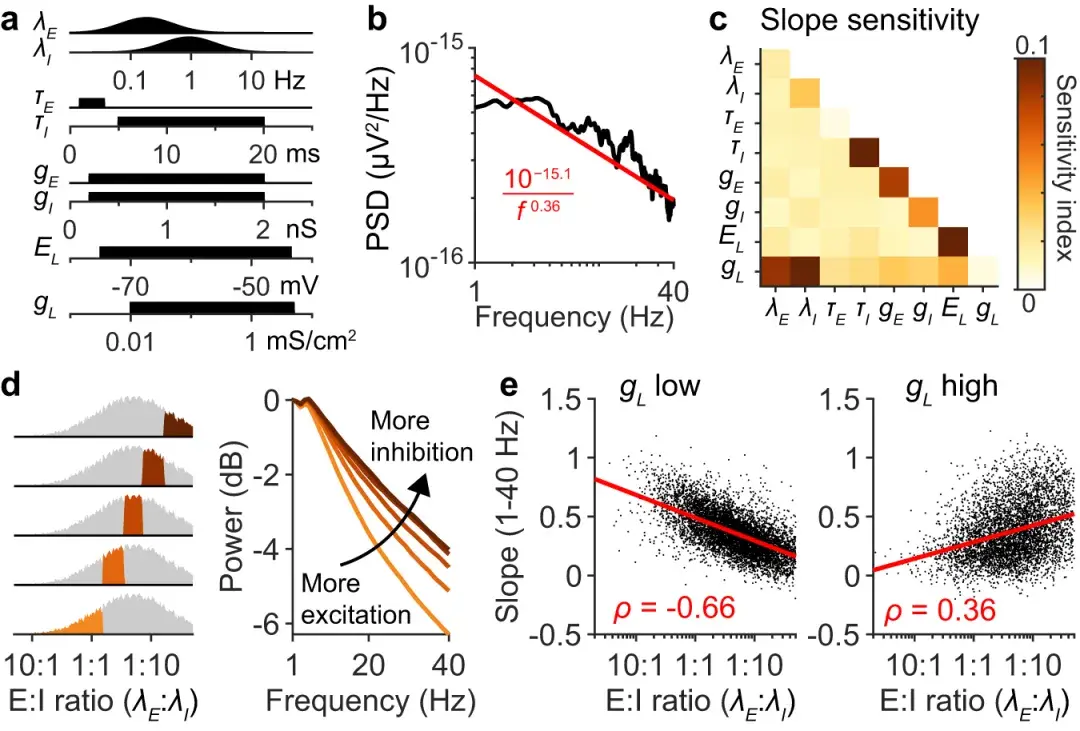
a :模型参数的抽样分布,与 图1g 中使用的相同 。
b :单神经元脑电图谱示例,用方程在 1 到 40 Hz 之间。
c :光谱斜率β对模型参数(具有一阶和二阶相互作用)的敏感性,根据 20,000 个模拟光谱计算得出。
d :根据和之间的比率对模拟光谱进行平均。较低的 E:I 比率对应于更多的抑制。
e :频谱斜率根据每次模拟的 E:I 比率绘制。左图:漏电导较低的模拟( mS cm − 2;n = 7366 次模拟)。右图:漏电导较高的模拟( mS cm −2;n = 5184 次模拟)。
a :接收完全亚临界输入的神经元的酉谱( 见图 5a )请注意,尽管突触输入动态没有变化,但单一谱的变化。b与 ( a ) 相同,但对于接收完全振荡输入的神经元 ( 见图 5a )。c拟合光谱趋势的示例。d来自 ( c )的光谱,通过将光谱除以各自的洛伦兹拟合来消除趋势。
e :引起的脑电图谱密度变化。
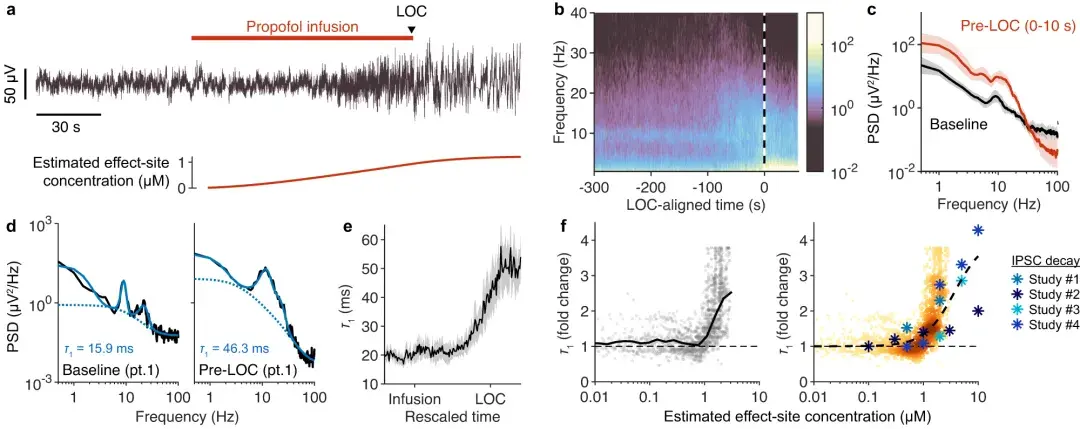
a :接受异丙酚输注直至意识丧失(LOC)的受试者的**性脑电图信号,Cz 记录位点。异丙酚效应部位的估计浓度如下图所示。
b :14 名受试者的平均 LOC 对齐频谱图。
c :基线处的平均功率谱(黑色),在输注异丙酚之前 0 到 10 秒之间取平均值;在输注异丙酚之后(红色),在 LOC 之前 0 到 10 秒之间取平均值。阴影反映平均值的 95% 置信区间(n = 14 名受试者)。
d :基线、丙泊酚输注前 0-10 秒(左,黑色)和 LOC 前 0-10 秒(右,黑色)的**性脑电图频谱。
e :参数,根据 2 秒窗口中计算的光谱拟合估计,相对于重新缩放的时间绘制。阴影反映平均值的 95% 置信区间(n = 14 名受试者)。
f :估计值的倍数变化相对于异丙酚估计效应位点浓度绘制。黑线标记异丙酚每个浓度的平均值。右:左图的估计剂量反应图(较深的颜色反映较高的点密度)。
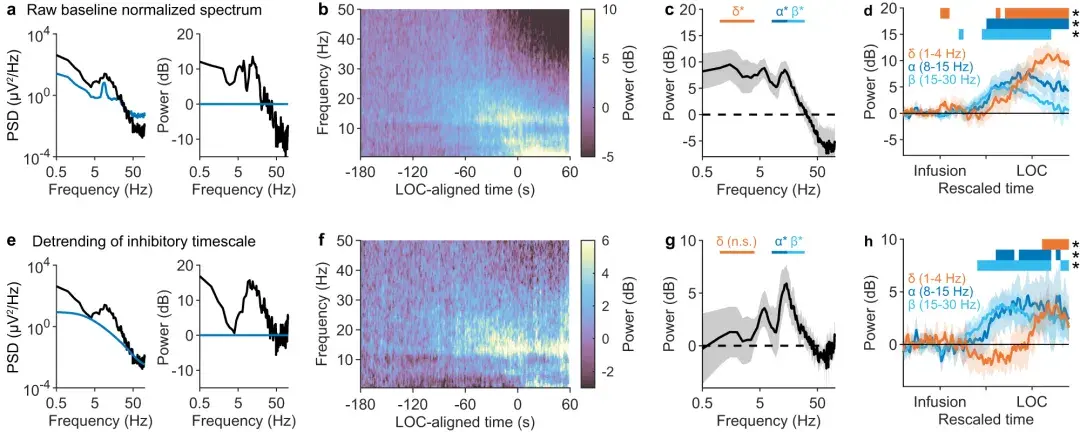
a 左 :单个受试者 LOC(黑色)后的**性脑电图功率谱,叠加在基线功率谱(蓝色)上。a 右:LOC(黑色)归一化至基线后的功率谱。
b :平均基线归一化频谱图(n = 14 名受试者)。
c :LOC 之前 0-10 秒的平均基线归一化功率(阴影:平均值的 95% 置信区间)。在 delta( p = 0.001,右尾符号检验;n = 14 名受试者)、alpha(p ≈ 10 −4)和 beta(p = 0.007)频段内,功率显着升高。
d :α、β 和 δ 功率的变化,与异丙酚输注时刻和 LOC 一致,在受试者之间进行平均(阴影:平均值的 95% 置信区间)。图表上方的条形表示重新调整时间的 0.05 长段 ,其中相应频带功率有统计上显着的增加(p < 0.05,右尾符号检验)。
e 左 :LOC 之后的 EEG 频谱,与a相同,叠加有拟合的抑制时间尺度。未显示拟合的高斯峰。e 右 :去趋势功率,定义为相对于拟合抑制时间尺度的功率,以分贝为单位。
f :平均去趋势频谱图,标准化至基线(n = 14 名受试者)。
g :LOC 之前 0-10 秒的去趋势功率,标准化至基线(阴影:平均值的 95% 置信区间)。α ( p = 0.006) 和 beta ( p = 0.001)频带内的功率显着升高 ,但 delta ( p = 0.40) 频带内的功率没有显着升高。显着性检验同( c )。
h :与 ( d ) 相同,但对于去趋势功效,标准化为基线。
非周期性神经活动可以对脑电图信号做出贡献,并且频谱趋势进一步受到许多生理机制的影响,例如兴奋/抑制平衡和突触时间尺度。
研究发现,光谱指数不**是大脑节律的综合测量,因此提供了大脑状态的补充生物标志物。然而,另一个发现中,谱指数没有单一的生理学解释。
**,研究得出的结论是,在量化大脑节律时,脑电图谱确实需要去趋势化,但前提是突触后电流特性被系统地改变。否则,去趋势可能会给脑节律量化带来重大错误,因此应该避免。
CONTACT 联系我们
联系人:樊女士
电话:18900616086 京显
邮箱:18900616086@163.com changxian-el@hotmail.com
地址:北京市海淀区中关村南大街5号二683号楼
联系方式▼ 更多咨询关注小程序▼


 产品展示
产品展示
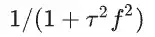

 http://bitbrain.cn/Zz_www.eastsummit.net/index.html
http://bitbrain.cn/Zz_www.eastsummit.net/index.html 
 销售一部
销售一部